双糖和多糖
双糖和多糖都是单糖分子通过分子间脱水后以苷键连接而成的化合物。
一、双糖
双糖广泛存在于自然界,它由两个相同的或不同的单糖单元构成。连接两个单糖的苷键有以下两种情况:
- 第一种情况,连接双糖的苷键是由两个单糖的半缩醛羟基脱水而成,因此分子中已没有半缩醛羟基,不能通过互变生成开链糖,也就没有还原性和变旋光现象,这种苷键连接的双糖为非还原性双糖。
- 第二种情况,一个单糖分子的半缩醛羟基与另一单糖分子中的醇型羟基之间脱水形成双糖,这种双糖分子中还有半缩醛羟基,因而有还原性和变旋光现象,为还原性双糖。
麦芽糖、纤维二糖、乳糖为还原糖,蔗糖为非还原糖。
单糖环状结构有α-和β-两种构型,这两种构型的半缩醛羟基都可参与苷键的形成,因此苷键就有α-苷键和β-苷键之分。
(一)麦芽糖
麦芽糖(maltose)存在于麦芽中,麦芽中的淀粉酶将淀粉水解而生成麦芽糖。此外,淀粉在稀酸中部分水解时可得麦芽糖。麦芽糖结晶含一分子结晶水,熔点103℃(分解),易溶于水,有变旋光现象,比旋光度为+136°。麦芽糖的结构如下:
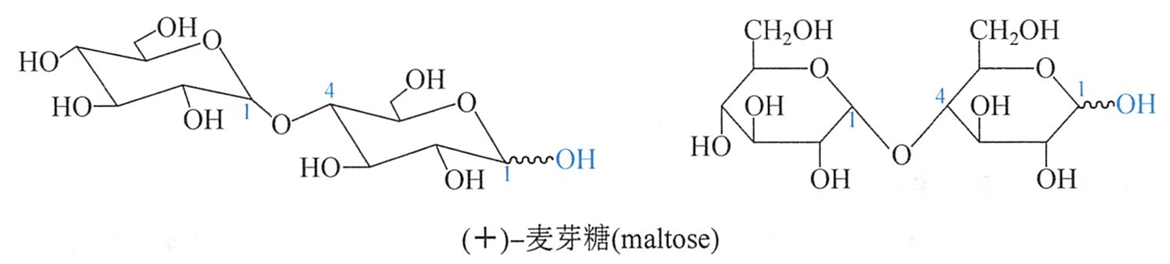
由上可知麦芽糖是由两分子D-葡萄糖以α-1,4-苷键连接构成的,成苷部分的葡萄糖是以吡喃环形式存在。麦芽糖分子结构中还有一个半缩醛羟基。麦芽糖是还原糖。
(二)纤维二糖
纤维二糖(cellobiose)是由纤维素部分水解得到。纤维二糖化学性质与麦芽糖相似,为还原糖,有变旋现象,水解后生成两分子D-葡萄糖。
与麦芽糖不同的是纤维二糖只能被β-葡萄糖苷酶水解,纤维二糖是以β-1,4-糖苷键组成的双糖,全名为4-O-(β-吡喃葡萄糖基)-D-吡喃葡萄糖。它的结构如下:
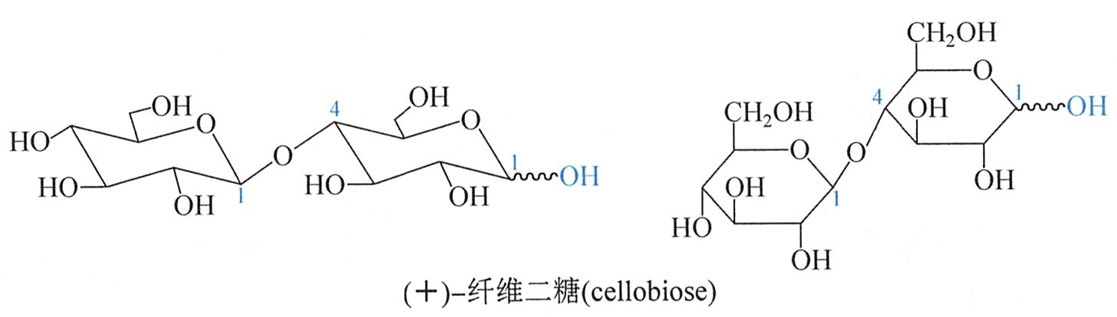
纤维二糖与麦芽糖虽只是苷键的构型不同,但生理上却有很大差别。麦芽糖有甜味,可在人体内分解消化,但纤维二糖既无甜味,也不能被人体消化吸收。
(三)乳糖
乳糖(lactose)存在于哺乳动物的乳汁中,人乳汁中乳糖含量为7%~8%,牛乳中含量为4%~ 5%。工业上可从制取奶酪的副产物(乳清)中获得乳糖。
乳糖也是还原糖,有变旋光现象。当用苦杏仁酶水解时,可得等量的D-半乳糖和D-葡萄糖,乳糖被溴水氧化后,水解可得到D-半乳糖和D-葡萄糖酸,故它是由半乳糖半缩醛羟基与D-葡萄糖的羟基键合而成。
根据苦杏仁酶专一性地水解β-糖苷键的特点及它的氧化、甲基化和水解反应得知,葡萄糖的C4羟基参与形成苷键。因此乳糖是β-1,4-糖苷键的双糖,其名称为4-O-(β-D-吡喃半乳糖基)-D-吡喃葡萄糖。其结构式为:
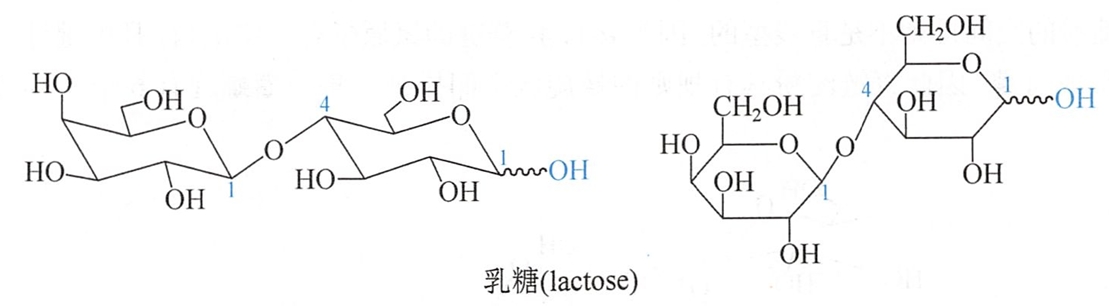
乳糖的结晶含一分子结晶水,熔点202℃,溶于水,比旋光度为+53.5°。医药上常利用其吸湿性小作为药物的稀释剂以配制散剂和片剂。
(四)蔗糖
蔗糖(sucrose))是自然界分布最广的双糖,尤其在甘蔗和甜菜中含量最高,故有蔗糖或甜菜糖之称。蔗糖被稀酸水解,产生等量的D-葡萄糖和D-果糖。
蔗糖没有还原性,也无变旋光作用,说明结构中已无半缩醛羟基。其苷键由葡萄糖的半缩醛羟基和果糖的半缩酮羟基脱水而成。蔗糖既可被α-葡萄糖苷酶水解也可被β-果糖苷酶水解生成相同产物,可知蔗糖既是α-D-葡萄糖苷也是β-D-果糖苷。后经X-衍射研究及全合成,确定了蔗糖为α-D-吡喃葡萄糖基-β-D-呋喃果糖苷,也可称为β-D-呋喃果糖基-α-D-吡喃葡萄糖苷。其结构如下:
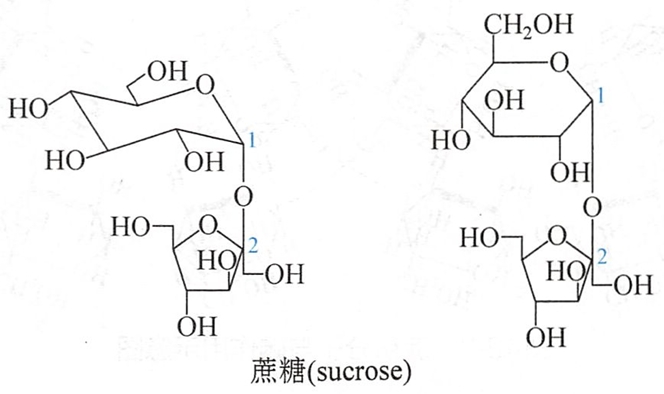
蔗糖是右旋糖,比旋光度为+66.7°,水解后生成等量的D-葡萄糖和D-果糖的混合物,其比旋光度为-19.7°,与水解前的旋光方向相反,因此把蔗糖的水解反应称为转化反应,水解后的混合物称为转化糖(invertsugar)。蜂蜜中大部分是转化糖。蜜蜂体内有一种能催化水解蔗糖的酶,这种酶被称为转化酶(invertase)。

二、多糖
多糖是由许多单糖分子以苷键相连形成的高分子化合物。如淀粉、纤维素、糖原。自然界大多数多糖含有80~100个单元的单糖。
多糖可以水解,但要经历多步过程,先生成相对分子质量较小的多糖,然后是寡糖,最后是单糖。
多糖大多数为无定形粉末,没有甜味。大多数不溶于水。个别多糖能与水形成胶体溶液。
(一)淀粉
淀粉(starch)广泛地分布于植物界,是人类获取糖类的主要源泉。淀粉是白色无定形粉末。它是由直链淀粉(amylose)和支链淀粉(amylopectin)两部分构成。
直链淀粉在淀粉中的含量约为20%,不易溶于冷水,在热水中有一定溶解度,分子量比支链淀粉少,一般由250~300个D-葡萄糖以α-1, 4苷键连接而成的直链化合物。
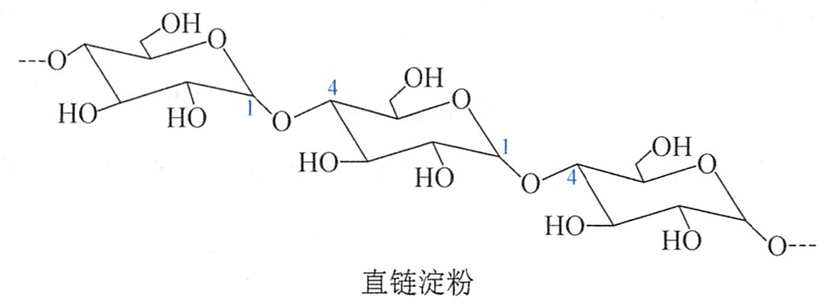
直链淀粉的空间排列不是直线型的,因为α-1,4-苷键的氧原子有一定键角,且单键可自由转动,羟基间可形成氢键,因此直链淀粉具有规则的螺旋状空间排列。每一圈螺旋有6个α-D-葡萄糖基(图16-2)。
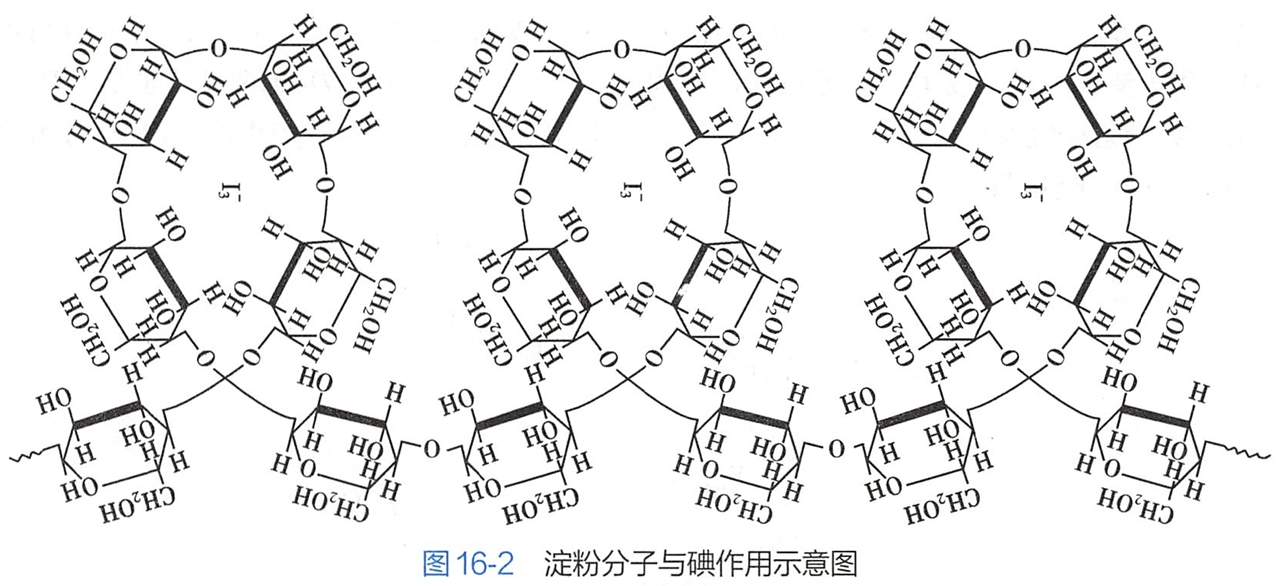
淀粉遇碘显蓝色,这是淀粉的定性鉴定反应。目前认为显色机制是碘分子钻入螺旋空隙中形成有色的复合物(图16-2)。
支链淀粉在淀粉中的含量约占80%,不溶于水中,与热水作用则膨胀成糊状。一般含有6000~40000个D-葡萄糖。在支链淀粉分子中,主链有α-1,4-苷键连接,而分枝处为β-1,6-苷键,结构如下:
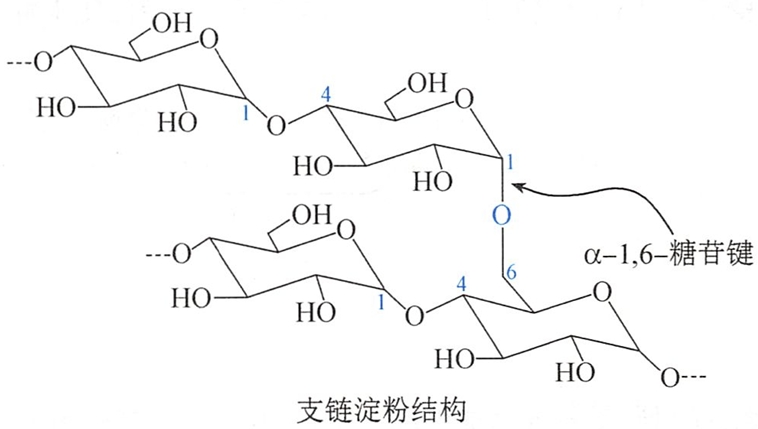
在支链淀粉分子的直链上,每隔20~25个D-葡萄糖单元就有一个以α-1,6-苷键连接的分支,因此其结构比直链淀粉复杂。支链淀粉可与碘生成紫红色的配合物。
淀粉在水解过程中可先生成糊精,它是相对分子质量比淀粉小的多糖,能溶于水,具有极强黏性。相对分子质量较大的糊精遇碘显红色,叫红糊精,再水解变成无色的糊精,无色糊精有还原性。淀粉在水解过程大致如下:
淀粉→红糊精→无色糊精→麦芽糖→葡萄糖
环糊精(cyclodextrin)是淀粉经环糊精糖基转化酶水解得到的多种环状低聚糖的总称。一般情况下环糊精是由六、七和八个D-(+)-葡萄糖通过α-1,4-糖苷键结合可分别得到α,β和γ三种环糊精。
(二)糖原
糖原(glycoge)是无色粉末,易溶于水,遇碘呈紫红色。
糖原主要存在于动物的肝脏和肌肉中,肝脏中糖原的含量达10%~20%,肌肉中的含量约4%。
其功能与植物淀粉相似,是葡萄糖的贮存形式。当血液中葡萄糖含量低于正常水平时,糖原即可分解为葡萄糖,供给机体能量。
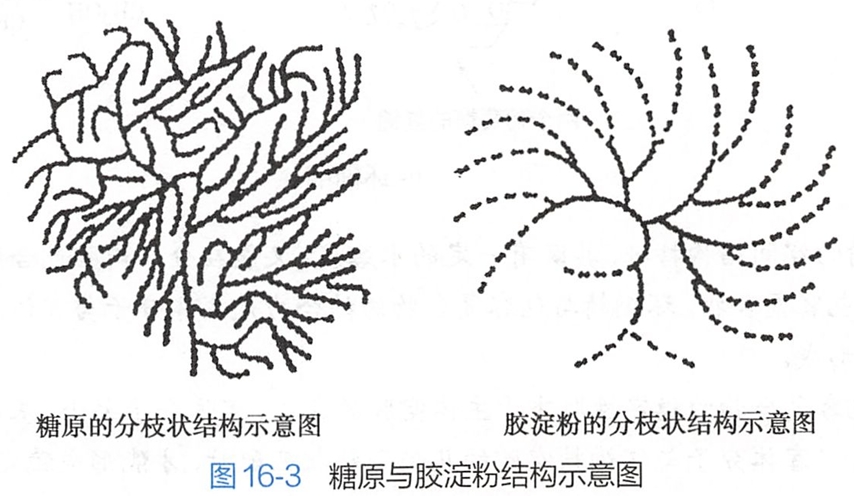
糖原的结构与支链淀粉相似,但分支更密,支链淀粉中每隔20~25个葡萄糖残基就出现一个α-1,6-苷键,而糖原只相隔8~10个葡萄糖残基就出现一个α-1,6-苷键(图16-3)。
(三)纤维素
纤维素(cellulose)是自然界最丰富的有机物。它是植物细胞壁的主要结构成分。植物千叶中含纤维素为10%~20%。木材中含纤维素50%,棉花含90%。纤维素是由D-葡萄糖以β-1,4-糖苷键结合的链状聚合物。在纤维素结构中不存在支链,分子链之间因氢键的作用而扭成绳索状。
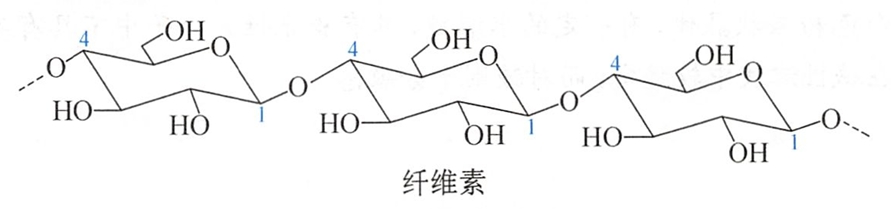
纤维素在盐酸水溶液中水解可得到D-葡萄糖。如用酶部分水解可产生纤维二糖。
纤维素虽然与淀粉一样由D-葡萄糖组成,但由于是以β-1,4糖苷键连接,不能被淀粉酶水解,因此人不能消化纤维素。但它可增强肠的蠕动,因此食入富含纤维素的食品有利于健康。食草动物的消化道中有一些微生物能分泌出可以水解β-1,4-糖苷的酶,可以消化纤维素。
纤维素无变旋现象,不易被氧化,但具有羟基一般反应,分子中游离的羟基经硝化和乙酰化后,可制成人造丝、火棉胶、电影胶片、硝基漆等。